Enigmes i certeses de la pesta, per Margarita Salvadó


Margarita Salvadó. Biòloga
Quan parlem de la pesta, sempre solem associar la malaltia al període anomenat pesta negra, que va assolar Europa entre els anys 1347 i 1353. Però al llarg de la història de la humanitat, la pesta ha estat present i continua estant-ne. Segons els butlletins d’informació de l’OMS, al segle XXI s’han produït brots de pesta a Madagascar (2015, 2014 i 2010), al Perú (2009), a la Xina (2006) i a la República Democràtica del Congo (2006 i 2005). Segons aquestes dades veiem que, actualment, la pesta es concentra a l’Amèrica Llatina, a l’Àfrica i a l’Àsia, tot i que també hi ha casos als Estats Units d’Amèrica, on se’n declaren entre 1 i 17, anualment, en alguns estats de l’oest, com Arizona, Oregon, Colorado i Califòrnia.
La pesta és una zoonosi, és a dir, una malaltia en què el bacteri es transmet a l’home a través dels rosegadors mitjançant un vector que és la puça. Els rosegadors infectats moren en grans quantitats i això provoca la migració de les puces des del seu reservori habitual fins als humans (hoste accidental), la picada de les quals penetra en l’organisme d’aquests últims (fig. 1). Distingim diferents tipus de pesta segons el lloc d’infecció. D’aquesta manera, parlem de pesta bubònica, en la qual la picada de la puça fa que els ganglis de la zona picada s’inflamin i es formin bubons. És la més comuna i no es propaga de persona a persona. També parlem de pesta septicèmica, quan el bacteri es dissemina a través del torrent sanguini, ja que la puça l’inocula directament. Aquesta no presenta bubons i se la coneix com a pesta negra, ja que quan els malats moren tenen els dits, el nas i orelles d’aquest color com a resposta de l’hoste infectat a les toxines del bacteri. El tercer tipus de pesta és l’anomenada pneumònica. Afecta els pulmons i el bacteri es transmet de persona a persona per les gotetes respiratòries de l’individu que van de l’infectat al què encara no ho està. Aquesta pesta és susceptible de ser utilitzada en terrorisme biològic (OMS 2016).
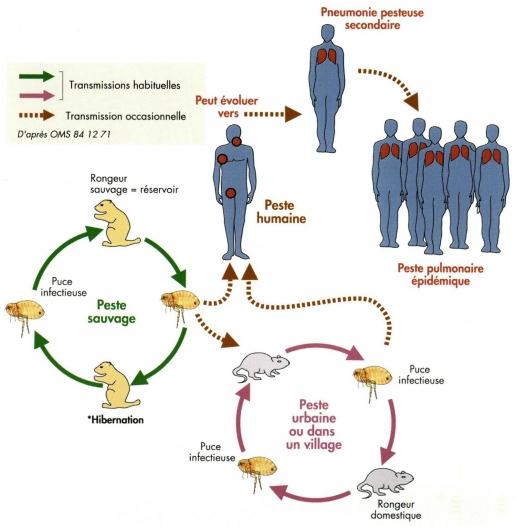
(Fig.1) Cicle de transmissió del bacteri Yersinia pestis. ATLAS DE LA PESTE à MADAGASCAR. Suzanne Chanteau. : http://www.openedition.org/6540
De les nombroses epidèmies que ha patit la història humana, les tres pandèmies que s’accepta que han estat causades per la pesta són, la pesta de Justinià (541-542 dC), la primera, que es va estendre d’Egipte a les àrees que envolten la Mediterrània. La segona va ser la coneguda com a pesta negra (1347-1353) i es va propagar des del Mar Caspi fins a gairebé tots els països d’Europa. Va causar la mort d’un terç de la població europea i va persistir en el continent fins al segle XVII, ja que va anar provocant brots successius fins que, progressivament, les ones epidèmiques van anar disminuint. La tercera pandèmia es va iniciar a la regió de Yunnan a la Xina, a la segona meitat del segle XIX, i es va estendre a escala mundial des de Hong Kong, en l’epidèmia de pesta del 1894. Durant aquesta última pandèmia, el bacteriòleg suís Alexandre Yersin va identificar el microorganisme o l’agent etiològic que n’era el causant, anomenat Yersinia pestis, que deu el nom al seu descobridor. Brots anteriors, com la pesta d’Atenes (430-427 aC) i la pesta Antonina (165-180 dC) també podien haver estat causades per la pesta però no hi ha cap evidència directa que permeti atribuir-la a Yersinia pestis (Drancourt, M. i Raoult D., 2002). Les conseqüències de les pandèmies de pesta han estat ben documentades i se sap que els diferents impactes demogràfics van ser espectaculars. La Pesta Negra va ser una de les pandèmies més devastadores de la història de la humanitat; s’estima que van morir entre 75 i 200 milions de persones a Euràsia. A Europa, va morir el 50-60% de la població entre 1347 i 1353; la població europea en aquella època era d’uns 80 milions. Alguns col·lapses econòmics i polítics del passat també han estat atribuïts, parcialment, als efectes devastadors de la pesta, com la de Justinià, que es creu que va tenir un paper important en el debilitament de l’Imperi bizantí. D’altres pestes anteriors s’han associat amb la pèrdua d’hegemonia de la Grècia clàssica i, és probable, també, que fos la causant del declivi de la força de l’exèrcit romà. (Little, L.K.[et al], 2007)
La pesta sempre ha estat un motiu d’estudi i de debat per part dels historiadors i dels científics. En els darrers anys se n’han publicat molts d’articles científics. Això es deu al fet que s’han aplicat tècniques genètiques a l’estudi de genomes antics de Yersinia pestis (microorganisme causal de la pesta), una pràctica que s’anomena paleogenètica o antropologia/paleontologia molecular, i que es defineix com la ciència que estudia la conformació molecular de l’ADN que s’ha trobat en fòssils de qualsevol tipus, i les relacions que té aquesta informació genètica amb l’ADN modern. Quan es tracta de buscar ADN de microorganismes causants de malaltia en esquelets humans, la metodologia que es fa servir és el desenterrament de les restes i la utilització de tècniques de radiocarboni per fer-ne la datació. Generalment, l’extracció de l’ADN del microorganisme es fa a partir de la polpa dentària, ja que la durabilitat, juntament amb l’esterilitat natural, fa que sigui un material adequat per basar aquestes investigacions. A l’ADN antic se l’anomena aADN. El perquè de l’interès de l’aplicació d’aquestes tècniques en la pandèmia de la pesta, és poder resoldre temes que són objecte de debat.
Entre aquests temes que han estat objecte de debat i els que encara ho són destaquen: 1.- el fet de si el mateix agent etiològic havia estat el causant de les tres pandèmies històriques; 2.- des d’on havia evolucionat aquest microorganisme amb la virulència amb què ho va fer; 3.- on s’havia establert el (els) reservori(s) del bacteri de la pesta a Europa en la segona pandèmia −la pesta negra−,que va continuar colpejant Europa durant quatre segles en brots posteriors que no van cessar fins a final del segle XVIII i que va desaparèixer de manera desconeguda (el focus de pesta selvàtica (pesta selvàtica és la que només afecta els rosegadors salvatges i es transmet entre ells mitjançant puces infectades) és avui present dia a gairebé tot el món però a Europa hi és absent, 4.- i si hi va haver una altra via de transmissió diferent de la que coneixem actualment, atesa la velocitat i l’eficàcia amb què es va propagar la segona pandèmia.
L’any 1998 es va detectar per primera vegada aADN de Yersinia pestis (Drancourt, M. .[et al],1998) a la polpa dental d’esquelets extrets de tombes franceses de persones mortes de pesta entre els segles XVI i XVIII. Aquest fet ha portat els científics a augmentar els esforços per detectar l’agent causant de la pesta en esquelets, tant de la segona pandèmia com de la primera. Fins no fa gaire, les etiologies de la pesta negra −en particular de la primera− van despertar no només controvèrsia entre els historiadors sinó també entre els científics. Alguns investigadors tenien dubtes sobre el fet que el bacteri Yersinia pestis hagués causat aquestes pandèmies, ja que més aviat pensaven que es tractava d’una malaltia viral. L’anàlisi molecular de l’aADN procedent de material arqueològic ha demostrat, de manera inequívoca, que Yersinia pestis ha tingut un paper important en les dues pandèmies històriques (Haensch, S., 2010; Schuenemann, V.J., 2011; Bos, K.I., 2011; Harbeck, M., 2013; Wagner, D.M., 2014).
L’any 2015, Simon Rasmussen i els seus col·laboradors van investigar l’origen de Yersinia pestis mitjançant la seqüenciació dels genomes de bacteris antics procedents de dents d’humans de l’edat del bronze a Europa i a l’Àsia (fig. 2). Aquestes troballes suggereixen que la soca virulenta de Yersinis pestis, transmesa per les puces que va causar les pandèmies històriques de pesta bubònica, va evolucionar a partir d’un llinatge seu menys patogen que va infectar poblacions humanes molt abans d’allò que evidencien els brots de pesta registrats. Van estudiar l’ aADN obtingut de les dents de 101 individus que van viure a l’edat del bronze a Europa i a l’Àsia (Allentoft[et al], 2015), i van trobar que set individus mostraven seqüències semblants a les de Yersinia pestis. Després de les investigacions fetes, van concloure que els estudis moleculars indicaven una ramificació ràpida de Yersinia pestis fa 5.783 anys (fig.3) i que havia evolucionat d’un bacteri menys virulent amb potencial zoonòtic, com Yersinia pseudotuberculosis. També van ser capaços d’obtenir una sèrie temporal de l’adquisició dels elements moleculars de patogenicitat, per convertir-lo en un dels bacteris més letals mai observats pels éssers humans. Així mateix van concloure, que soques de l’ancestre de Yersinia pestis eren presents al final del quart mil·lenni abans de Crist i que es van difondre àmpliament a través d’Euràsia, pel cap baix des de principis del mil·lenni següent. L’aparició de la pesta en individus euroasiàtics de l’edat del bronze indica que les infeccions de pesta eren comunes, almenys, 3.000 anys abans de les dades registrades històricament. S’ha arribat a suggerir que aquestes primeres pestes podien haver estat les responsables de la disminució de la població entre finals del quart mil·lenni abans de Crist i principis del tercer (Hinz, M.[et al]., 2012, Shennan, S.[et al], 2013).
Amb aquestes dades, els dos primers temes debatuts en el transcurs de la història queden tancats definitivament. A continuació veurem què és el que ens diu la ciència pel que fa als altres dos que queden.
 (Fig. 2) Situació geològica de les ubicacions arqueològiques de Yersinia pestis a l’edat del bronze (S. Rasmussen [et al.], 2015).
(Fig. 2) Situació geològica de les ubicacions arqueològiques de Yersinia pestis a l’edat del bronze (S. Rasmussen [et al.], 2015).
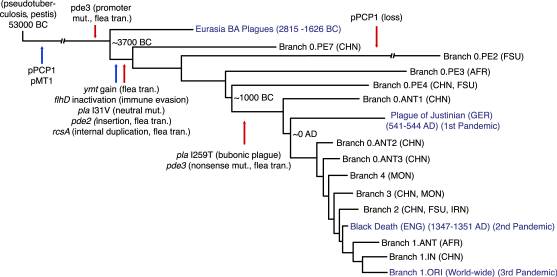
(Fig. 3) Evolució esquemàtica de Yersinia pestis des del seu ancestre Yersinia pseudotuberculosis on es veu com va adquirir els gens de virulència i les diferents branques filogenètiques trobades fins ara(S. Rasmussen [et al], 2015).
Un altre gran tema de discussió sempre ha estat quin va ser el reservori de la pesta durant la segona pandèmia, i perquè va desaparèixer sense deixar rastre al segle XVIII. Generalment s’assumeix que el bacteri va arribar de l’Àsia a Europa a través de les rutes comercials de la seda, que es va establir en reservoris de rosegadors urbans (Ell, Sr., 1984,Keeling, M.J., 2000) donat, a més, que l’Imperi Otomà va boicotejar el comerç amb l’oest i va tancar les rutes l’any 1453. Per tant, es creu que a partir d’aquests reservoris, la malaltia hauria causat brots successius en humans fins que el bacteri va desaparèixer. L’any 2015, Boris V. Schmid, de la Universitat d’Oslo, va dur a terme un estudi dirigit pel doctor Nils Christian Stenseth que tenia com a objectiu localitzar els reservoris de la pesta a Europa i a l’Àsia, basant-se en l’estudi de les fluctuacions climàtiques que precedeixen les epidèmies regionals. Els autors van fer servir 7.711 brots de pesta històrics i 15 cronologies d’anells dels arbres, com a registres substitutoris del clima donada l’estreta relació entre el clima i la formació d’anells de fusta (dendroclimatologia), i sota la condició que, igual que molts focus de pesta contemporània (Ben-Ari, T. [et al], 2011) els reservoris de l’època medieval també van ser sensibles a les fluctuacions climàtiques.
La hipòtesi climàtica de l’estudi, es basa en el coneixement que es té avui dia de la dinàmica que ha seguit la pesta en rosegadors silvestres i els efectes col·laterals que tenen lloc en éssers humans, i que es coneixen prou bé gràcies a algun focus de pesta actual esdevingut al centre de l’Àsia (Samia, N.I. [et al],2011). Per esmentar només un cas, la prevalença de pesta en el rosegador anomenat jerbu petit (Gerbillinae) al Kazakhstan està afectada fortament en sentit positiu quan hi ha primaveres més càlides i estius més humits, favorables al creixement de la vegetació,(Stenseth, N.C.[et al], 2006) i es correlaciona amb la densitat de població de jerbus petits i de puces (Reijniers, J.[et al], 2012). Quan el clima facilita una alta densitat de rosegadors també afavoreix l’extensió de la pesta a tota la regió (Davis, S.[et al], 2008), i quan la població de jerbus petits es col·lapsa en resposta als canvis climàtics la densitat de puces per jerbus s’incrementa, dramàticament, la qual cosa facilita encara més la propagació de la pesta a través de la població de rosegadors (Reijniers, J.[et al], 2014) provocant que les puces busquin hostes alternatius, inclosos éssers humans i animals domèstics. Coneixent el patró dels anells dels arbres actuals de l’Àsia que ens indiquen clima favorable a la pesta, Boris V. Schmid va estudiar les fluctuacions climàtiques que van precedir epidèmies de pesta regionals per localitzar els reservoris de pesta durant l’època medieval. Ho va fer basant-se en 7.711dades georeferenciats de brots històrics de pesta procedents d’anells d’arbres d’Europa i de l’Àsia. Dels 4.119 brots seleccionats, es van excloure els brots que probablement podrien haver estat causats per raons diferents en els reservoris locals com per exemple, els brots secundaris d’un brot inicial. En aquesta configuració força estricta, 24 brots que van tenir lloc en 19 ciutats es van associats, possiblement, a reservoris de pesta en vida silvestre. Es va documentar que vuit d’aquests 24 brots van ser conseqüència d’importacions marítimes de pesta des d’altres ciutats (Devaux, C.A., 2013), amb la qual cosa només van quedar 16 brots interessants per estudiar. L’anàlisi detallada del clima de la regió on van succeir aquests 16 brots a Europa no va revelar cap patró significatiu que pogués estar associat amb un reservori sensible al clima de la pesta i a la fauna local. Els resultats obtinguts van donar suport a un escenari en què, en el registre dels arbres de la regió del Karakorum i d’una zona més gran en l’època medieval, les fluctuacions del clima eren favorables perquè es poguessin promoure i sincronitzar els brots de pesta entre les poblacions de rosegadors de l’Àsia central (Davis, S. [et al], 2008). Quan el clima es tornava posteriorment menys favorable, facilitava el col·lapse de les poblacions de rosegadors infectats de pesta i obligava les puces a trobar hostes alternatius (Samia N.I. [et al], 2011). Els autors diuen que la gran quantitat de brots de pesta en vida silvestre a l’Àsia seria, durant el temps de la segona pandèmia de pesta, el reservori des d’on va anar arribant la pesta als ports d’Europa amb un retard d’uns 15 anys (fig. 4).
(Fig.4)Esquema del procés de reintroducció de la pesta a Europa. Una nova reintroducció de la pesta consta de tres etapes: la caiguda de la població de rosegadors 1 o 2 anys després de l’inici de la disminució de les condicions climàtiques; el transport de la malaltia a Europa al cap de 10 o 12 anys, i la seva propagació a través de la xarxa europea de comerç marítim en menys de 3 anys. També es mostren les rutes comercials (marró i línies blaves), els reservoris de pesta en vida silvestre coneguda (taronja) i els ports marítims europeus (cercles grisos) associats a la nova introducció marítima (Schmich, B.V., 2015).
Els autors conclouen que aquest estudi ofereix una perspectiva diferent de la de transmissió a llarga distància de Yersinia pestis a través d’Euràsia i desplaça la teoria d’una única introducció en el moment de l’inici de la segona pandèmia a una altra d’entrada intermitent impulsada per les condicions climàtiques i com, en cada entrada, arribarien noves soques procedents dels reservoris establerts a l’Àsia. Si aquesta perspectiva és correcta, en l’anàlisi d’aADN de Yersinia pestis de víctimes de la pesta de la segona pandèmia hi hauria diversitat genètica (Bos K.I.[et al], 2011, Haensch, S. [et al]., 2010).
L’any 2016, Maria A. Spyrou i col·laboradors seus van publicar un estudi on es plantejaven aquesta qüestió i van dissenyar un estudi d’aADN antic de diferents desenterraments d’esquelets de morts a causa de la pesta. Van recollir mostres d’una fossa comuna a Barcelona, d’una tomba a la ciutat de Bólgar (Rússia) i d’una fossa comuna a Ellwangen (Alemanya). Per estimar-ne o confirmar-ne el període històric es va fer la datació dels fragments d’ossos i de les arrels de les dents amb radiocarboni. Les dates obtingudes van ser els anys 1300-1420 per a Barcelona, 1298-1388 per a Bólgari 1486-1627 per a Ellwangen. Les ciutats que es van fer servir en aquest estudi es van triar estratègicament per poder estudiar la ruta que va fer la soca de Yersinia pestis en aquesta pandèmia, per veure’n l’evolució genètica i si entre els genomes estudiats de diferents èpoques hi havia molta diversitat genètica.
Els resultats suggereixen que hi va haver una diversitat limitada de Yersinia pestis durant la fase primerenca de la pesta negra, cosa que fa pensar en una sola entrada de pesta a Europa; que posteriorment hi va haver una onada de pesta que va viatjar cap a l’est després de la pesta negra, que més tard va donar lloc a la pandèmia del segle XIX, i que hi va haver una participació del mateix llinatge de pesta en dos brots europeus després de la pesta negra amb 200 anys de diferència, les epidèmies de Marsella i d’Ellwangen.
En un altre estudi fet també l’any 2016, Lisa Seifert i els seus col·laboradors van estudiar l’aADN d’esquelets extrets d’una fossa comuna del segle XIV d’Alemanya i d’una altra del segle XVII, i van trobar que els genomes identificats en els dos desenterraments eren idèntics malgrat els 300 anys de diferència i aposten per les reintroduccions de Yersinia. pestis des del centre d’Àsia en múltiples onades durant la segona pandèmia, i per la persistència de Yersinia pestis a Europa en un hoste reservori encara desconegut. L’any 2010 en canvi, Stephanie Haensch i el seu equip van estudiar els genomes d’esquelets de fosses comunes del nord, del centre i del sud d’Alemanya, i van trobar dos llinatges diferents, un d’idèntic al de l’estudi fet per Seifert i l’altre de diferent, i els autors es van inclinar a pensar que la pesta va entrar a Europa per diferents entrades tal com suggereix l’estudi d’Schmid.
Caldria fer moltes més excavacions arqueològiques a tot Europa per comprovar molts més genomes de Yersinia pestis i veure si la seva diversitat és limitada i procedeixen d’un mateix llinatge, o bé si apareixen molts llinatges que certifiquin que hi va haver només una entrada o moltes, s’hi ha molta diversitat.
Finalment l’any 2012 el grup de treball de Marsella, liderat per Michael Drancourt, va proposar la hipòtesi que la transmissió de la pesta es va transmetre a través dels polls, és a dir, que el factor clau en l’explosió d’epidèmies de la pesta negra va poder ser la transmissió directa d’humà a humà després de l’adaptació de la Yersinia pestis a un vector alternatiu com és el poll del cos.
Agraïments: a la Dra Emilia Gutíerrez, professora Titular d’Ecologia, Facultat de Biologia, Universitat de Barcelona, per les seves aportacions i revisió d’aquest manuscrit.
Referències
Allentoft, M.E.;Sikora, M.;Sjögren, K.G.; Rasmussen, S.; Rasmussen, M.;Stenderup, J.;Damgaard, P.B.; Schroeder, H.;Ahlström, T.;Vinner, L. «Population genomics of Bronze Age Eurasia».Nature, 2015;522:167–172. PMID: 26062507.DOI:1038/nature14507
Ben-Ari, T.[et al]. «Plague and climate: Scales matter». PLoSPathog, 2011; (9):e1002160
Bos, K.I.;Schuenemann, V.J.; Golding, G.B.;Burbano, H.A.;Waglechner, N.;Coombes, B.K.[et al]. «A draft genome of Yersinia pestis from victims of the Black Death». Nature, 2011; 478:506–510. DOI:10.1038/nature10549. PMID: 21993626
Chanteau, Suzanne. Atlas de la peste à Madagascar. Nouvelle édition [en ligne]. Marseille : IRD Éditions, http://books.openedition.org/irdeditions/6582>. ISBN : 9782709918510. DOI : 10.4000/books.irdeditions.6582
Davis, S.;Trapman, P.;Leirs, H;Begon, M.;Heesterbeek, J.A.P. «The abundance threshold or plague as a critical percolation phenomenon». Nature, 2008; 54 (7204):634–637
Devaux, C.A. «Small oversights that led to the Great Plague of Marseille (1720-1723): Lessons from the past». Infect Genet Evol, 2013; 14:169–185
Drancourt,M.;Aboudharam, G.;Signoli, M.;Dutour, O.;Raoult, D. «Detection of 400-year-old Yersinia pestis DNA in human dental pulp: An approach to the diagnosis of ancient septicemia». Proc. Natl. Acad. Sci., USA. 1998; 95: 12637–12640. PMID: 9770538
Drancourt, M.;Raoult, D. «Molecular insights into the history of plague».Microbes Infect, 2002;4:105–109. [PubMed: 11825781]
M. Drancourt. Plague in the genomic area. Clin Microbiol Infect 2012; 18: 224–230
Ell, Sr. «Immunity as a factor in the epidemiology of medieval plague». Rev. Infect. Dis., 1984; 6(6):866–879
Haensch, S.; Bianucci, R.;Signoli, M.;Rajerison, M.; Schultz, M.;Kacki, S.[et al]. «Distinct Clones of Yersinia pestis Caused the Black Death». PLoSPathog, 2010; 6(10):e1001134. DOI: 10.1371/journal.ppat. 1001134. PMID: 20949072
Harbeck, M.; Seifert, L.;Hänsch, S.; Wagner, D.M.;Birdsell, D; Parise, K.L.[et al]. «Yersinia pestis DNA from Skeletal Remains from the 6th Century AD Reveals Insights into Justinianic Plague». PLoSPathog, 2013; 9 (5):e1003349. DOI: 10.1371/journal.ppat.1003349. PMID: 23658525
Hinz, M.;Feeser, I.;Sjögren, K.G.; Müller, J. «Demography and the intensity of cultural activities: an evaluation of Funnel Beaker Societies (4200–2800 cal BC)»Archaeol. Sci, 2012;39:3331–3340.
Keeling, M.J.; Gilligan, C.A.«Metapopulation dynamics of bubonic plague». Nature, 2000;407(6806):903–906.
Little, L.K.; Hays, J.N.;Morony, M.; Kennedy, H.N.;Stathakopoulos, D.; Sarris, P.;Stoclet, A.J.;Kulikowski, M.;Maddicott, J.; Dooley, A. «Plague and theend of antiquity: The pandemic of 541–750».Cambridge University Press, 2007.
OMS 2016. http://www.who.int/mediacentre/factsheets/fs267/es/
Rasmussen, S.;Allentoft, M.E.; Nielsen, K.; Orlando, L.;Sikora, M.;Sjören, K.G.[et al]. «Early Divergent Strains of Yersinia pestis in Eurasia 5,000 Years Ago». Cell. 2015; 22:571–82. doi: 1016/j.cell.2015.10.009
Reijniers, J.;Begon, M.;Ageyev, V.S.;Leirs, H.«Plague epizootic cycles in Central Asia». Biol Lett, 2014; 10(6):pii: 20140302
Reijniers, J[et al]. «A curve of thresholds governs plague epizootics in Central Asia». Ecol Lett, 2012; 15(6):554–560.
Samia, N.I., [et al]. «Dynamics of the plague-wildlife-human system in Central Asia are controlled by two epidemiological thresholds». Proc. Natl. Acad. Sci. USA, 2011; 08(35):14527–14532.
Schmid, B.V.;Büntgen, U.;Easterday, W.R.;Ginzler, C.;Walløe, L.;Bramanti, B.[et al]. «Climate-driven introduction of the Black Death and successive plague reintroductions into Europe». Proc. Natl. Acad. Sci. USA, 2015; 112(10):3020–3025. DOI: 10.1073/pnas.1412887112. PMID: 25713390
Schuenemann, V.J.;Bos, K.;DeWitte, S.;Schmedes, S.; Jamieson, J.;Mittnik, A.[et al]. «Targeted enrichment of ancient pathogens yielding the pPCP1 plasmid of Yersinia pestis from victims of the Black Death». Natl.Acad.Sci. USA, 2011; (108)38:E746–E752. DOI:10.1073/pnas.1105107108. PMID: 21876176
Seifert, L.;Wiechmann, I.;Harbeck, M.; Thomas, A.;Grupe, G.;Projahn, M.[et al]. (2016) «GenotypingYersinia pestisin Historical Plague: Evidence for Long-Term Persistence of Y. pestis in Europe from the 14th to the 17th Century». PLoS ONE 11(1): e0145194. DOI:10.1371/journal.pone.0145194
Shennan, S.; Downey, S.S.; Timpson, A.;Edinborough, K.;Colledge, S.;Kerig, T.; Manning, K.; Thomas, M.G. «Regional population collapse followed initial agriculture booms in mid-Holocene Europe». Commun.,2013;4:2486
25. Spyrou, M.A.; Tukhbatova, R.I.; Feldman, M.; Drath, J.; Kacki, S.; Beltrán de Heredia, J.; Arnold, S.; Sitdikov, A.G.; Castex, D.; Wahl, J.; Gazimzyanov, I.R.; Nurgaliev, D.K.; Herbig, A.; Bos, K.I.; Krause, J.«Historical Y. pestis Genomes Reveal the European Black Death as the Source of Ancient and Modern Plague Pandemics». Cell Host Microbe, 2016.19(6):874-81. DOI: 10.1016/j.chom.2016.05.012. PMID: 27281573
Stenseth, N.C., [et al]. «Plague dynamics are driven by climate variation». Proc. Natl. Acad.Sci. USA, 2006. 103(35):13110–13115
Wagner, D.M.;Klunk, J.;Harbeck, M.;Devault, A.;Waglechner, N.;Sahl, J.W.[et al]. «Yersinia pestis and the Plague of Justinian 541–543 AD: a genomic analysis». Lancet Infect Dis., 2014. 14:319–326. DOI: 10. 1016/S1473-3099(13)70323-2. PMID: 24480148




